
.
livello difficile
.
ARGOMENTO: BIOLOGIA MARINA
PERIODO: ODIERNO
AREA: na
parole chiave: balene, energetica, fisiologia
Abbiamo, con grande interesse, esaminato lo studio intitolato “Estimating energetics in cetaceans from respiratory frequency: why we need to understand physiology” di Fahlman et al. (2016), e abbiamo apprezzato la loro enfasi sulla necessità di comprendere la fisiologia quando vengono stimati i tassi metabolici di campo (FMR) dei cetacei. In questo documento, Fahlman et al. hanno utilizzato i dati da loro raccolti su delfini bottlenose in cattività (Tursiops truncatus), sia a riposo che dopo un periodo di nuoto in superficie, per testare la validità dell’uso di variabili respiratorie di cetacei misticeti di grandi dimensioni che vivono liberi per la stima del loro FMR ( Field Metabolic Rates).

Questo approccio fu usato per la prima volta nella balena blu (Balaenoptera musculus) dal premio Nobel August Krogh, (Krogh, 1934), e finora è stato l’unico modo per ottenere stime utili del dispendio energetico nei grandi cetacei in libertà. Il metodo sfrutta il fatto che i mammiferi marini sono di fatto dei subacquei che trattengono il respiro e che traggono vantaggio dall’ottimizzare il loro tempo sott’acqua conducendo uno scambio respiratorio durante brevi periodi di superficie, durante i quali vengono scambiati rapidamente grandi volumi d’aria (Blix e Folkow, 1995).
L’approccio si basa sull’equazione:
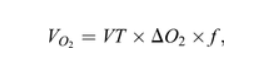
dove VO2 è il tasso di assorbimento di ossigeno (l min-1); VT è il volume corrente (l breath-1); ΔO2 è la differenza nella concentrazione di ossigeno tra inspirata (O2 in) ed aria espirata (O2 ex); ΔO2 = O2in-O2ex (%); e f è la frequenza respiratoria (respiro min-1).
Nella loro analisi, Fahlman et al. hanno valutato tre metodi, tutti basati su Eqn 1 (metodi A, B e C).
In particolare, nel metodo A sono stati usati i dati di Armstrong e Siegfried (1991) e Dolphin (1987), e in B i nostri dati (Blix e Folkow, 1995), in entrambi i casi per stimare l’FMR nei grandi misticeti; mentre nel metodo C, Fahlman et al. hanno usato i dati dai loro delfini. In tutti i casi i risultati sono stati confrontati con le misurazioni dirette di Fahlman et al. del VO2 negli stessi delfini. Così facendo, non sorprende che il metodo C abbia funzionato bene, mentre i metodi A e B hanno dimostrato di sovrastimare il valore “vero” di circa il 200-500%. In estrema sintesi questi sono i risultati che, anche se con qualche riserva, Fahlman et al. offrono sulla validità del loro confronto. Tuttavia è necessario sottolineare che Fahlman et al. hanno condotto i loro raffronti attraverso diversi livelli di attività, subordine e più di 20 ordini di grandezza rispetto alla massa corporea.

In questo modo, hanno scelto di applicare relazioni allometriche empiriche per la predizione dei volumi polmonari invece di usare i valori misurati sui loro delfini. Il fatto che i risultati dei diversi metodi differiscano sostanzialmente non dovrebbe quindi sorprendere più di quanto l’uso della formula di Du Bois, usata per la stima della superficie nell’uomo (Du Bois e Du Bois, 1916) si dimostrerebbe inadeguata per il calcolo della superficie, per esempio, nelle giraffe. Inoltre, se Fahlman et al. avessero stimato i volumi polmonari dei loro delfini usando l’equazione empirica di Wahrenbrock et alii (1974) per le balene grigie (Eschrichtius robustus), avrebbero ottenuto valori negativi.
Per illustrare ulteriormente il punto, è stata testata la validità del Metodo C di Fahlman et al., usando i dati esistenti per un grande misticeto. Wahrenbrock et al. (1974) hanno misurato che un giovane di balena grigia di 4000 kg a riposo ha un VO2 corrispondente a circa 2 × il metabolismo basale di Kleiber (BMR), cioè come previsto per qualsiasi mammifero immaturo e in crescita (Kleiber, 1961).

Usando il Metodo C con i dati di Fahlman et al. per VT a riposo (32% della capacità polmonare totale) e ΔO2 (4,94%), e Wahrenbrock e alii (1974) avendo misurato i valori del volume polmonare (236 litri) e la frequenza respiratoria a riposo ( ~1 breath min-1) per un vitello di 4000 kg, si ottiene un VO2 di 3,7 litri min-1, che è inferiore al 35% del VO2 di 10,7 litri min-1 misurato.

Se applichiamo il metodo C di Fahlman ai dati (Blix and Folkow, 1995), otteniamo che una balena minke (Balaenoptera acutorostrata) sarebbe in grado di nuotare a 2-3 ms-1 in un FMR, valore che è del 20% inferiore al BMR di Kleiber nei mammiferi in generale (Kleiber, 1961), o per buona misura, ad un FMR simile (in W kg-0,75) come nel bradipo tridattilo (Bradypus variegatus) (Nagy e Montgomery, 1980).
In sintesi gli autori ritengono che ”One would assume that, with proper physiological insight, the extraordinarily large discrepancies between the outcomes of the different methods would have inspired similar exercises, and that Fahlman et al.’s paper, in consequence, would not have been published in its present form.“
Link all’articolo originale da Biology Open 2017 6: 306 doi: 10.1242 / bio.023929
.
Riferimenti
– Armstrong, A. J. and Siegfried, W. R. (1991). Consumption of Antarctic krill by minke whales. Antarctic Sci. 3, 13-18. doi:10.1017/S0954102091000044 CrossRefGoogle Scholar
– Blix, A. S. and Folkow, L. P. (1995). Daily energy expenditure in free living minke whales. Acta Physiol. Scand. 153, 61-66. doi:10.1111/j.1748-1716.1995.tb09834.x CrossRefPubMedGoogle Scholar
– Dolphin, W. F. (1987). Dive behavior and estimated energy expenditure of foraging humpback whales in southeast Alaska. Can. J. Zool. 65, 354-362. doi:10.1139/z87-055CrossRefGoogle Scholar
– Du Bois, D. and Du Bois, E. F. (1916). A formula to estimate the approximate surface area if height and weight be known. Arch. Intern. Med. 17, 863-871. doi:10.1001/archinte.1916.00080130010002 CrossRefPubMedGoogle Scholar
– Fahlman, A., van der Hoop, J., Moore, M. J., Levine, G., Rocho-Levine, J. and Brodsky, M. (2016). Estimating energetics in cetaceans from respiratory frequency: why we need to understand physiology. Biol. Open 5, 436-442. doi:10.1242/bio.017251 Abstract/FREE Full TextGoogle Scholar
– Kleiber, M. (1961). The Fire of Life – An Introduction to Animal Energetics. New York: John Wiley & Sons, Inc..Google Scholar
– Krogh, A. (1934). Physiology of the blue whale. Nature 133, 635-637. doi:10.1038/133635a0CrossRefGoogle Scholar
– Nagy, K. A. and Montgomery, G. G. (1980). Field metabolic rate, water flux, and food consumption in three-toed sloths (Bradypus variegatus). J. Mammal. 61, 465-472. doi:10.2307/1379840 Abstract/FREE Full TextGoogle Scholar
– Wahrenbrock, E. A., Maruschak, G. F., Elsner, R. and Kenny, D. W. (1974). Respiration and metabolism in two baleen whale calves. Mar. Fish. Rev. 36, 3-8.Google Scholar
Una sorpresa per te su Amazon Music unlimited Scopri i vantaggi di Amazon Prime
Alcune delle foto presenti in questo blog possono essere state prese dal web, citandone ove possibile gli autori e/o le fonti. Se qualcuno desiderasse specificarne l’autore o rimuoverle, può scrivere a infoocean4future@gmail.com e provvederemo immediatamente alla correzione dell’articolo
,
–
Footnotes
The authors declare no competing or financial interests. © 2017. Published by The Company of Biologists Ltd http://creativecommons.org/licenses/by/3.0. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0), which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.

- autore
- ultimi articoli
è composta da oltre 60 collaboratori che lavorano in smart working, selezionati tra esperti di settore di diverse discipline. Hanno il compito di selezionare argomenti di particolare interesse, redigendo articoli basati su studi recenti. I contenuti degli stessi restano di responsabilità degli autori che sono ovviamente sempre citati. Eventuali quesiti possono essere inviati alla Redazione (infoocean4future@gmail.com) che, quando possibile, provvederà ad inoltrarli agli Autori.